Chapter
4 The Neural Bases of Language
In this chapter, the author
attempts to provide some current view on the nature of the neural bases of
human language. His focus is on the
cortical-striatal-cortical circuits which yield the reiterative
ability. The faculty not only
plays a critical role in human speech and confers the flexibility
of human thought processes but probably derives from elements of neural
circuits that regulate motor control.
Besides, he also discuss studies that are exploring the neural bases of
the brain��s dictionary, neural
plasticity, and the lateralization of
the brain.
I.
Neuroanatomy
(1.)
Introduction
A.
The brain is divided in two cerebral hemispheres, along the anterior to
posterior axis.
B.
Structures closer to the midline of the brain are ��medial,�� and
those out from it are ��lateral.��
C.
Neocortex has neurons arranged in six layers.
(a)
paleocortex
(b)
basal ganglia
(c)
thalamus
(d)
hippocampus
(e)
cerebellum
(f)
other subcortical structures in the basal ganglia and other subcortical
structures are represented right and left.
D.
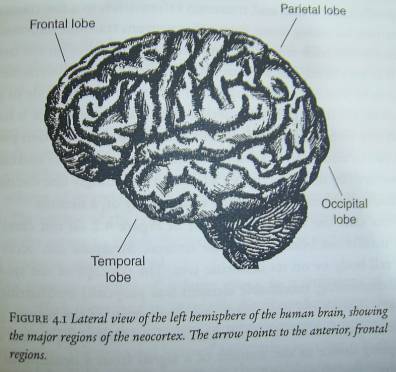 The
surface of the brain is shown in the lateral view:
The
surface of the brain is shown in the lateral view:
(2.)
Neurons
A.
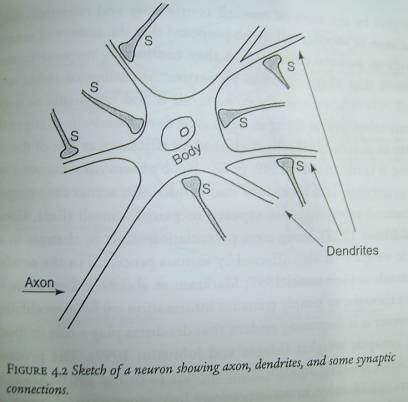
Neurons connect to each other and transmit information by means of
dendrites and axons.
B.
A cluster of dendrites is associated with each neuron.
C.
Synapses are structures that determine the degree to which an incoming
signal will cause the cell body to generate an electrical pulse, the action
potential or spike that is transmits out without decrement on its axon.
D.
The synapse acts as a coupling device in the transfer of a signal from a
dendrite to the neuron.
(3.)
Distributed Neural Networks
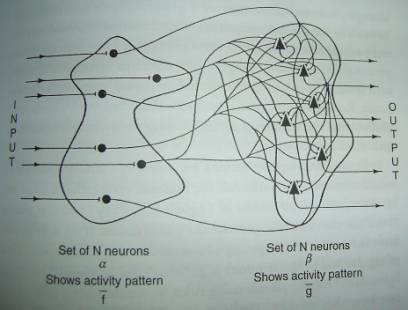
A.
Every neuron in set�\projects to every
neuron in set�].
B.
This drawing, where N=6, grossly understates the size and connectivity of
the nervous system and single neurons and single synapses have little effect on
the discharge patterns of the group as a whole.
(4.)
Understanding Distributed Neural Networks
A.
Large sections of the network can be destroyed without losing memory
traces.
B.
When the cortical areas are destroyed, people do not lose their
preexisting language or their native dialect.
C.
Transient loss of language may occur, but after a period of months, a
person��s premorbid linguistic ability returns.
II.
Electrical Power Grids: A Metaphor for Distributed Networks
(1.)
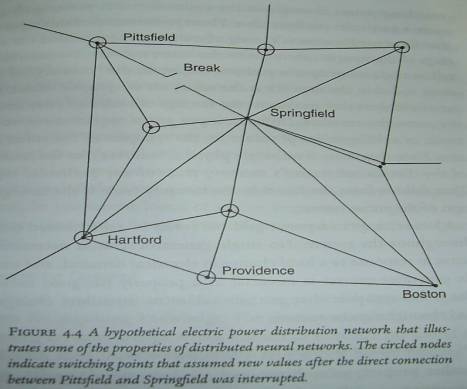
Electrical Power Grids:
(2.)
The Robustness of Distributed Neural Networks
A.
The robust nature of memory trace follows from the fact that information
is coded by the synaptic settings of the entire system.
B.
A distributed network also can be perturbed by events taking place far
away from them.
C.
If switches fail to correct the problem in time, an isolated event that
overloads a switching point can propagate through the network.
D.
If the network is properly designed and monitored, a single power plant
failure or downed transmission line would not interrupt your electrical supply.
E.
The memory traces that code the motor pattern generators that result in
speech, the cognitive pattern generators specifying the rules of syntax, and the
lexicon clearly have widely distributed cortical representations.
(3.)
Associative Learning
A.
Imitation and associative learning are powerful ��general�� procedures
for acquiring knowledge��
Pavlov��s dogs
B.
Associative learning has been demonstrated in Aplysia californius ��
a gastropod mollusk.
C.
Associative learning has a selective value to any animal because it
allows rapid phenotypical changes in response to new environmental conditions.
III.
Cortical Architecture�w�wCortical
Maps
(1.)
The human brain is not a unitary distributed neural network.
(2.)
Different parts of the cortex have different cytoarchitectonic
structures, in other words, different distributions of neurons in the layers
that form the cortex.
(3.)
The cytoarchitectural maps of brains made by Brodmann generally were
thought to reflect discrete functional distinctions. (It is becoming clear that the situation is more complex than
Brodmann thought.)
(4.)
Broca��s and adjacent cortical areas also are implicated in manual motor
control.
(5.)
Cortical areas and subcortical structures carry out different local
operations and play a part in circuits regulating different aspects of behavior.
(6.)
It clearly is not a ��module�� dedicated to language and language
alone.
(7.)
The feed-forward and back connections that typify all cortical areas
manifest themselves in Broca��s area being activated during speech perception
and sentence comprehension.
(8.)
Many areas of cortex are malleable and can take in new functions due to
damage to the brain or birth defects.
IV.
The Neurophysiologic Toolkit
(1.)
Direct Electrophysiologic Recording
A.
Direct observation of neuronal activity in the brain of a living animal
involves the placement of microelectrodes.
B.
These electrodes conduct electrical signals induced by the
electrochemical communications between neurons.
(2.)
Tracer studies
A.
One of the properties of axons is that amino acids, the building blocks
of proteins, are transmitted from the cell body down the length of an axon.
B.
All traditional tracer techniques involve injecting substances into the
brain of a living animal, waiting for the tracer to be transported, and then
sacrificing the animal.
C.
A recent advance in tracer techniques employs a variation of MRI
technology that allows neural circuits to be traced in living human subjects.
(3.)
MRI, fMRI, and PET Imaging
A.
Functional magnetic resonance imaging (FMRI) is the most recent
technique, and it is a variant on the structural magnetic resonance imaging (MRI)
systems that are routinely used for imaging brain structures.
B.
FMRI can map out the flow and transport of deoxygenated blood (the BOLD
signal) that is an indirect marker of metabolic activity, hence neural activity,
in the brain.
C.
Event-relater FMRI involves close synchrony of tasks with the FMRI BOLD
signal, permitting investigation of the time course of neural activity.
D.
Positron emission tomography (PET) involves injecting a short-half life
radioactive tracer into the bloodstream.
(4.)
CT Scans
Computerized
tomography (CT) show slices of brain anatomy reconstructed by computer
processing of multiple X-ray exposures taken at different angles and planes.
(5.)
ERP
Event-related
potentials (ERPs) can reveal transient electrical activity recorded by means of
electrodes placed in contact with a person��s scalp. The technique involves recording by means of the procedures
commonly used to record EEG brain activity in clinical settings.
V.
Brain�w�wBehavior
Models
(1.)
Phrenology
(2.)
Modular Theories
VI.
Neural Circuit Models

(1.)
Neurophysiologic activity must be considered at two levels.
First, distinct neuroanatomic structures usually do not regulate an
observable behavior.
A.
The neuronal population that carries out the local operation in a given
part of the brain projects to anatomically segregated neuronal populations in
other neuroanatomic structures.
B.
Circuits linking anatomically segregated populations of neurons form
neural systems carrying out processes in different parts of the brain. The
systems are the neural bases of complex behaviors.
(2.)
Second, different regions of the neocortex and different subcortical
structures are specialized to process particular stimuli, visual or auditory,
while other regions perform specific operations that regulate aspects of motor
control or holding information in short-term memory. The local operations form a set of neural computations that
link together in complex neural circuits.
(3.)
The neural mechanism that carries out the instruction set manifested in
my pecking at my computer��s keyboard is a circuit, linking neuronal
populations in different neuroanatomic structures in many parts of the brain.
VII.
Experiments in Nature
(1.)
The traditional Broca-Wernicke Model
A.
Although the Broca-Wernicke model has the virtue of simplicity, it is
wrong. Clinical evidence shows that permanent loss language does not occur
without subcortical damage even when Broca��s or Wernicke��s areas have been
destroyed.
B.
Damage to subcortical structures, sparing the cortex, can produce aphasic
syndromes.
C.
Language impairments occurred that ranged from fairly mild disorders in
the patient��s ability to recall words to ��global aphasia�� in which the
patient produced very limited nonpropositional speech.
D.
Damage to the internal capsule (the nerve fibers that connect neocortex
to subcortical structures0 and basal ganglia structures, the putamen, and
caudate nucleus resulted in impaired speech production similar to that of the
classic aphasias, as well other cognitive deficits.)
(2.)
The Basal Ganglia and Cortical�w�wStriatal
Circuits
A.
Basal ganglia dysfunction can produce the constellation of motor, syntax,
and cognitive deficits associated with Broca��s syndrome.
B.
The basal ganglia are subcortical structures located deep within the
brain. They are primitive neural structures that can be traced back to anurans.
C.
The basal ganglia in humans and other primates include the caudate
nucleus and the lentiform nucleus, which constitute the striatum.
VIII.
Neurodegenerative Diseases
Damage to
striatal and associated subcortical structures that support a cortical-striatal-cotrical
circuit can result in a behavioral deficit that was thought to derive from
damage to the region if the cortex served by the circuit.
The primary
deficits of these neurodegenerative diseases are motoric; tremors, rigidity, and
repeated movement patterns occur. However,
these subcortical diseases also cause linguistic and cognitive deficits.
(1.)
Probable Basal Ganglia Operations
The
functional basal ganglia complex includes the putamen, caudate nucleus, globus
pallidus, and close connections to the substantia nigra, thalamus, and other
structures. The system essentially
acts as a sequencing engine. Within
the basal ganglia, information is transmitted by inhibitory and excitatory
channels.
(2.)
Neuroimaging Studies�w�wSet
Shifting and Sequencing
(3.)
Implicit Associative Learning
(4.)
Blobology
A.
This procedure attempts to identify neural structures that are key
factors in regulating a particular aspect of behavior by subtracting the pattern
of activity pattern associates with the experimental stimuli.
B.
Blobology overlooks the fact that circuits linking activity in many
neuroanatomic structures generally carry out complex, observable behaviors.
C.
The neuroanatomic structures implicated in listening to acoustic signals
must necessarily be activated when we interpret word.
And there is no guarantee that the part of the brain that supposedly is
the seat of word comprehension is not also activated in some other behavior that
was not explored in the particular experiment.
IX.
Evidence for Basal Ganglia Activity in Cortical�wStriatal�wCortical
Circuits
(1.)
Broca��s Syndrome
(2.)
Speech Sequencing
X.
Linguistic Deficits of Broca��s Syndrome
Contrary to the traditional
Broca��-Wernicke model that assigns speech production to Broca��s area,
higher-level linguistic deficits also occur in Broca��s aphasic syndrome. They consistently show reduced auditory semantic priming
effects, especially when the acoustic signal is degraded.
Cognitive Deficits of Broca��s Syndrome
XI.
Parkinson��s Disease (PD)
(1.)
VOT Sequencing Deficits
(2.)
Sentence Comprehension Deficits
(3.)
Cognitive Set-Shifting Deficits
(4.)
Cognitive Planning
XII.
Mount Everest as an Experiment in Nature�w�wHypoxia
XIII.
Cerebellum
XIV.
Verbal Working Memory
XV.
Dynamic Systems: Throwing More Resources at the Problem
XVI.
The Brain��s dictionary
The Dog Who
Did Not Bark: When Procedural Knowledge Becomes Declarative Knowledge
XVII.
Cortical
Plasticity
XVIII.
Basal Ganglia Grammar
Motor
Control Programs and the Rules of Syntax
XIX.
Brain Lateralization and Language